Brunobian
Usuario
Virus encontrados en el río Cam, en Cambridge, podrían convertirse en la próxima generación de antibióticos, de acuerdo con lo anunciado en la Universidad de Edinburgo, en el Reino Unido. Con la actual sobre-prescripción de antibióticos para el tratamiento de infecciones bacterianas, y pacientes que no completan propiamente sus tratamientos, muchas bacterias son capaces de conformar genes para resistir a los antibióticos simplemente intercambiando material genético entre si. El MRSA (Staphylococcus aureus resistente a Meticilina), la cepa resistente a múltiples drogas del Staphylococcus Aureus, y las nuevas cepas emergentes de Clostridium dificile, han forzado a los investigadores a buscar un abordaje completamente diferente para combatir estas bacterias. Bacteriófago: Esquema (fuente: Wikipedia)Bacteriófago: Esquema (fuente: Wikipedia)"Utilizando un virus que solo ataca bacterias, llamado bacteriófago - y algunos bacteriófagos solo atacan tipos específicos de bacterias - podemos tratar infecciones apuntando a la cepa exacta de la bacteria causante de la enfermedad", dice Ana Toribio, del Wellcome Trust Sanger Institute en Hinxton, Cambridgeshire, Reino Unido. "Esto es mucho más específico que la terapia convencional con antibióticos" Los científicos utilizaron un pariente cercano de Escherichia coli, la bacteria que comúnmente causa intoxicación e infecciones gastrointestinales en humanos, llamado Citrobacter rodentium, que tiene exactamente el mismo efecto gastrointestinal en ratones. Ellos fueron capaces de tratar los ratones infectados con un cóctel de bacteriófagos obtenidos en el río Cam que ataca al Citrobacter rodentium. Actualmente los investigadores están optimizando la selección de los virus mediante análisis de ADN para utilizar los bacteriófagos con diferentes perfiles. Ataque de bacteriófagos: visualización 3D (fuente: Wikipedia)Ataque de bacteriófagos: visualización 3D (fuente: Wikipedia)"El uso de bacteriófagos en vez del tradicional tratamiento con antibióticos de amplio espectro, que esencialmente tratan de matar todas las bacterias que se les cruzan, es mucho mejor porque no alteran el balance microbial en el cuerpo", dice el Dr. Derek Pickard del Wellcome Trust Sanger Institute. "Todos necesitamos de buenas bacterias para ayudarnos a combatir infecciones, para digerir nuestros alimentos y proveernos de nutrientes esenciales, y los antibióticos convencionales pueden matar también esas bacterias, mientras luchan contra las que provocan la enfermedad". Hasta hace poco, los tratamientos basados en bacteriófagos han sido ignorados en Europa Occidental y Estados Unidos. Los principales reportes clínicos en humanos han sido producidos en Europa Oriental, particularmente en el Instituto Bacteriofago Tbilisi en Georgia, donde los bacteriófagos han sido utilizado con éxito en el tratamiento de infecciones tales como úlceras y llagas en diabéticos. "Cuanto más podamos desarrollar el tratamiento y comprender los obstáculos encontrados en el uso de este método para tratar infecciones intestinales, mayor es la probabilidad de maximizar su posibilidad de éxito en el largo plazo", dice Ana Toribio. "Hemos encontrado que el uso de una variedad de bacteriófagos (en vez de solo uno) para tratar una enfermedad tiene muchos beneficios en el ataque de una cepa peligrosa de bacteria, sobrepasando cualquier resistencia potencial al bacteriófago mediante mutaciones de la bacteria." Fuente Traducida: http://www.ciencianet.com.ar/154/nuevos-virus-para-tratar-enfermedades-bacteriales Fuente Original: http://www.physorg.com/news108006098.html
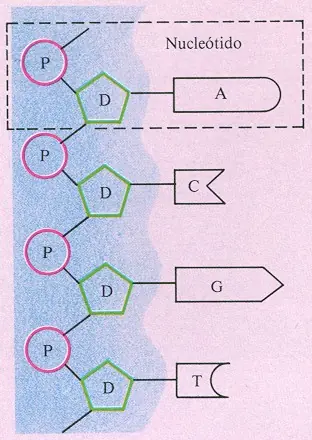
Bueno aca les dejo un poco de info de la revista ciencia hoy, espero que les sirva Adn: una molécula maravillosa Néstor O. Bianchi Instituto Multidisciplinario de Biología Celular CIC - CONICET Hasta 1944 no se sospechaba que el ácido disoxirribonucleico, ADN, fuera la molécula capaz de asegurar la transmisión de los caracteres hereditarios de célula a célula, generación tras generación. Su limitada variedad química no permitía suponer que poseyera la versatilidad y ductilidad necesarias para almacenar la información genética de los seres vivos. No fue entonces sin asombro que a partir de ese año el ADN se convirtió en centro de interés de la biología. Hoy sabemos que esta molécula, capaz de autoduplicarse y transmitir así su información, es una estructura dinámica y cambiante. Los avances logrados en el estudio de sus formas auguran un tiempo en el que se pueda comprender mejor su arquitectura y topología y la manera en que los microcambios moleculares provocan macrocambios en el funcionamiento genético. Quizá todo comenzó cuando alguno de nuestros primitivos antecesores, descansando tras un duro día de caza, observaba distraídamente un conjunto de guanacos o de caballos salvajes y consideraba el hecho de que de una pareja de caballos sólo nacen caballos y de una pareja de guanacos sólo nacen guanacos.... El reconocimiento de la herencia ha de haber sido, muy probablemente, una de las primeras ideas científicas aprehendidas por el hombre. Sin embargo, hasta después de pasada la primera mitad de nuestro siglo no se sabía con certeza dónde se almacenaba ni cómo se transmitía de célula a célula y del individuo a su descendencia la información hereditaria. Han pasado más de treinta años desde que J.D. Watson y F.H. Crick, eligiendo los datos más relevantes de un cúmulo de información y jugando con recortes de cartón y modelos de alambre y metal, fueron capaces de develar la estructura de la doble hélice de la molécula del ácido desoxirribonucleico, ADN, y formularon los principios de almacenamiento y transmisión de la información hereditaria. Este hallazgo les valió el premio Nobel, que compartieron con M.H.F. Wilkins. Los libros The Double Helix de Watson y The Eight Day of Creation de Freeland Judson describen en detalle la historia del descubrimiento, la personalidad de sus protagonistas, el sabor amargo de las frustraciones y la alegría del éxito. El develado de la estructura del ADN se debió más al ingenio que al trabajo; fue el triunfo de la cigarra sobre la hormiga. Fue, sobre todo, el comienzo de una apasionante historia que llega a nuestros días poblada aún de sorpresas y problemas por resolver. En la presente revisión nos referiremos a algunas de las particularidades estructurales de esta maravillosa molécula a la que las primeras décadas de nuestro siglo, aun cuando se conocía ya entonces su vinculación con los cromosomas y genes, consideraron demasiado simple como para otorgarle una función escencial en la transmisión de los caracteres hereditarios. La macromolécula de ADN está constituida por dos cadenas de nucleótidos complementarias. Como puede verse en la figura 1, los nucleótidos, que están formados por la unión de un grupo fosfato (ácido fosfórico), un azúcar (la molécula pentosa 2-desoxi-D-ribosa) y una base nitrogenada, se encadenan entre sí mediante la unión del azúcar de uno de ellos con el azúcar del contiguo a través del fosfato. El grupo fosfato y la desoxirribosa constituyen una suerte de columna vertebral que sirve de sostén a bases nitrogenadas de cuatro tipos diferentes; dos de ellas -la adenina (A) y la guanina(G)- son púricas, con estructura en doble anillo, las otras dos -la citosina(C) y la timina(T)- son pirimídicas, con estructura en anillo simple. La figura 2 muestra cómo se organizan los componentes químicos de cuatro eslabones de una molécula de ADN y al mismo tiempo presenta la estructura de las cuatro bases nitrogenadas; éstas se orientan hacia las bases de la cadena complementaria con la que se ligan por medio de puentes de hidrógeno. La estructura de las bases es de tal naturaleza que la G sólo se une con la C (lo hace por medio de tres puentes hidrogenados) y la A con la T (lo hace por medio de dos de tales puentes). Esto significa que mientras la secuencia de bases de una de las cadenas no responde, en principio, a ningún orden preestablecido, el orden de las bases de la segunda estará fatalmente determinado por la primera. Al diseñar las figuras 1 y 2, que sólo presentan eslabones de una cadena, hemos elegido libremente el orden de las bases, pero si el lector deseara construir ahora los eslabones correspondientes de la otra cadena, advertiría el carácter obligatorio de la secuencia de bases. Esta obligatoriedad constituye elf undamento del sistema de transmisión de la información genética. En efecto, la duplicación del ADN supone, primero, la separación de las dos cadenas mediante mecanismos enzimáticos y luego, a causa del carácter obligatorio o complementario del apareamiento de bases, la síntesis enzimática de una cadena idéntica a aquella de la que se separó. Esta fidelidad de la duplicación asegura que la información genética contenida en esa molécula de ADN se transmita fielmente, sin cambios, de célula a célula, generación tras generación. Fig.l. Representación esquemática de cuatro eslabones de una cadena de nucleótidos. Cada nucleótido consta de un grupo fosfato (P),un azúcar (D) y alguna de las cuatro bases nitrogenadas, adenina (A), citosina (C), guanina (G) y timina (T). Fig. 2.Distribución de los componentes químicos de la cadena esquematizada en la figura 1. El grupo fosfato (rojo) y el azúcar (verde) constituyen la "columna vertebral" de la macromolécula (sombreado celeste). Los fosfatos ligan el carbono 5' de la desoxirribosa de un nucelótido con el carbono 3' de la adyacente. La información genética se codifica a través de secuencia de las cuatro bases nitrogenadas que se muestran en la figura (negro). El sombreado amarillo indica los puentes de hidrógeno que unirán dos cadenas complementarias. Las cadenas poliméricas complementarias se presentan como una doble hélice o espiral determinada por dos hebras -las columnas vertebrales constituidas por azúcares y fosfatos- que se enrollan en forma paralela alrededor de un eje imaginario a la manera de una escalera caracol. Esta disposición se denomina plectonémica. Escalones de la metafórica escalera serían los pares de bases unidas por puentes de hidrógeno (véase la figura 3). En el caso del ADN más frecuente, el de tipo B, el ancho de la doble hélice es de 20 Å (Å = Angstrom = 1/10.000.000 mm). Cada vuelta entera de la hélice tiene una longitud de 34 Å y la distancia entre dos pares de nucleótidos sucesivos es de 3,4 Å, de manera que se necesitan 10 pares de bases para completar un giro completo de la hélice. Asimismo, el sentido de giro se corresponde con el de las agujas del reloj (sentido horario). Como puede verse en la figura 2, los nucleótidos se unen mediante los fosfatos que conectan el carbono en la posición 5' de una pentosa con el carbono en la posición 3' de la adyacente. Estas uniones 5'-3' determinan la direccionalidad de las cadenas. La figura 3 nos permite advertir con claridad que la dirección del extremo 5' al extremo 3' de las dos cadenas complementarias es opuesta, antipolar o antiparalela. Puede observarse también la presencia de un surco mayor y un surco menor formados por los giros de la espiral plectonémica. El ADN presenta tres organizaciones espaciales diferentes, todas ellas con forma de hélice, a las que se ha denominado A, B y Z. La figura 4 muestra, en el caso de las bases complementarias G-C, por dónde pasa el eje imaginario de la hélice en cada uno de estos tres tipos de ADN. Como puede observarse, la ubicación del eje modifica la profundidad de los surcos mayor y menor. Esta misma figura nos ilustra acerca de cómo pueden situarse las pentosas de manera que los carbonos en posición 4' 'y 5' se orienten hacia el surco mayor (forma anti) o hacia el surco menor (forma syn). Estas orientaciones caracterizan a los distintos tipos de ADN, pues mientras las pentosas de las formas A y B se ubican según una modalidad anti, la forma Z presenta a la pentosa de la base C en posición anti y a la de la base G en posición syn, hecho que, como luego veremos, determina su forma peculiar. Por lo general, el ADN se presenta bajo la forma B, pero ya Watson y Crick observaron que en condiciones de moderada deshidratación podía adoptar la organización espacial A. En este caso se requieren 11 pares de bases para la ejecución de un giro completo de la hélice, lo cual determina una forma ligeramente "subenrollada" con respecto a la que presenta el tipo B. Asimismo, como indica la figura 4, debido al desplazamiento de los pares de bases en relación al eje longitudinal de la hélice, el tipo A manifiesta una marcada diferencia entre sus surcos (el mayor es mucho más profundo que el menor), distinción que en el tipo B no resulta particularmente notable. El ADN de tipo Z, del que luego nos ocuparemos en detalle, se diferencia más de las formas A y B que éstas entre sí. Watson y Crick describieron el modelo de las formas A y B del ADN mediante la interpretación de fotografías de difracción de rayos X obtenidas de fibras de ADN nativo, es decir, extraído de células. Sin embargo, la magnitud de la información que puede obtenerse de este tipo de estudios es limitada, debido a que las fibras de ADN son extremadamente largas, no cristalizan y habitualmente presentan un cierto desorden estructural. El método, a lo sumo, permite establecer la estructura de un ADN promedio o estadístico, el cual no necesariamente representa la estructura de las diferentes regiones de ADN A o B existentes en la naturaleza. Cualquier variación local de estructura que pueda resultar de una secuencia particular de bases, pasará inadvertida al analizar la difracción de rayos X inducida por una larga fibra de ADN. Resulta claro, entonces, que los ADN A y B de Watson y Crick son macromoléculas "abstractas" que representan el promedio de un conjunto de variantes moleculares que se suceden a lo largo de la cadena. El análisis estructural de estas variantes sólo ha sido posible en los últimos años, gracias al advenimiento de métodos eficaces para sintetizar cadenas muy cortas de ADN que se denominan oligómeros. Estas cadenas poseen entre 4 y 24 pares de bases cuya secuencia ha sido predeterminada por el investigador. Los oligómeros cristalizan con facilidad y, por lo tanto, cuando se los somete a difracción de rayos X, proveen una información que permite la reconstrucción espacial fidedigna de su estructura. Si además se desea saber cómo sería la arquitectura de una larga fibra de ADN formada por un gran número de repeticiones del oligómero, pueden suministrarse los datos moleculares del mismo a una computadora, la cual se encarga de simular un modelo espacial con tantas repeticiones como se le ordene. A fines de la década del 70 se iniciaron las investigaciones destinadas a develar la arquitectura de distintas secuencias de ADN de los tipos A y B. Así, en1979, Wang sintetizó y analizó el hexámero CGCGCG y Drew el tetrámero CGCG (la notación convencional de una secuencia de bases de ADN de doble cadena consiste en citar las bases de una cadena leída del extremo 5' al extremo 3'). Con gran sorpresa pudo observarse que ninguno de estos oligómeros mostraba una estructura acorde con las dos formas conocidas de ADN. En efecto, la arquitectura de estas secuencias mostraba un giro de la doble hélice en sentido antihorario, y una peculiar conformación en zig-zag de las cadenas de fosfato-azúcar. Debido a este recorrido zigzagueante, la nueva forma recibió el nombre de ADN Z (véase la figura 5). Una de las características relevantes del polímero Z es la conformación de sus surcos mayor y menor: el primero prácticamente no existe, el segundo es profundo y cavernoso. Esta configuración es exactamente opuesta a la del ADN A, y si observamos una vez más la figura 4 advertiremos la razón: los pares de bases de uno y otro se encuentran desplazados en relación a los ejes de las respectivas hélices de manera opuesta. La síntesis artificial de nuevos oligómeros permitió obtener las ya conocidas formas A y B del ADN y demostró que la forma Z sólo aparece cuando existen secuencias de pares de bases donde alternan las C con las G ó viceversa. Como ya señalamos y vimos, además, en la figura 4, las pentosas están habitualmente unidas a las bases en la forma anti, con sus carbonos 4' y 5' incluidos en el surco mayor. La base G tiene la particularidad de presentar tanto la forma anti como la forma syn de unión con la pentosa; en este último caso los carbonos 4' y 5' están incluidos en el surco menor. Así, en una secuencia CGCG...(o viceversa) en la que las pentosas de la guanina se hallen en posición syn, se producirá una alternancia de formas anti y syn en la columna vertebral de pentosas-fosfatos que dará lugar a la configuración en zig-zag y al enrollamiento antihorario de la doble hélice, es decir, a la forma Z. El descubrimiento del ADN Z fue inesperado y se debió al uso de oligómeros sintéticos para el estudio de la biofísica del ADN. La síntesis de nuevas secuencias cortas de ADN permitió obtener las ya conocidas formas A y B y determinar la magnitud de la variación estructural entre las moléculas "estadísticas" de Watson y Crick y éstas a las que bien podemos denominar "realistas". En efecto, estos tetrámeros ,hexámeros ,dodecámeros y demás oligómeros obtenidos artificialmente, reproducen formas existentes en la célula viva e ilustran el hecho de que a lo largo de la cadena de ADN de un cromosoma pueden existir importantes variaciones de conformación que producen una microheterogeneidad topográfica. Hemos de considerar, entonces, las diferencias que pueden existir entre el ADN promedio o estadístico de Watson y Crick y el ADN real. La rotación de la doble cadena de ADN implica la existencia de un giro entre dos pares de bases sucesivos. Se forma así un ángulo de giro (véase la figura 6) que es de 36° para la forma B de la molécula promedio de Watson y Crick y de 32,7° para la forma A dado que, como sabemos, se necesitan respectivamente 10 y 11 pares de bases para completar una vuelta, es decir 360°, de la doble cadena. Los estudios con oligómeros cristalizados de ADN muestran, sin embargo, que el ángulo de giro puede variar de 28° a 42° en la forma B y de 16° a 44° en la forma A. Estos rangos de variación son considerables y muestran hasta qué punto puede modificarse la estructura del ADN a lo largo de la molécula. Dado que la forma Z está enrollada en sentido antihorario,a los ángulos de giro entre dos pares de bases sucesivos se les adjudica signo negativo: para el ADN Z formado por secuencias GC, los ángulos de giro observados varían entre -52,9° y -49,7°, mientras que en el caso de secuencias CG lo hacen entre -9,6° y -7,4°. Con respecto a la distancia entre dos pares de nucleótidos sucesivos cabe señalar que se han registrado importantes variaciones; por ejemplo en el caso de la forma B dicha distancia varía hasta un 13 % alrededor del valor promedio de 3,4 Å que se ha indicado anteriormente. Otro de los parámetros biofísicos variable es el ángulo de alabeo. En todo par de bases complementarias, éstas se encuentran giradas una con respecto de la otra a lo largo del eje longitudinal del par. El ángulo de alabeo es el que resulta de este giro (véase la figura 7) y registra variaciones en los rangos de 9,2° a 21,6°, de 6,9° a 16,5° y de 1,6° a 7,2° para las formas A, B y Z respectivamente. Las consideraciones anteriores ilustran el grado de heterogeneidad estructural que puede existir a lo largo de una cadena de ADN. Además de los parámetros biofísicos aquí considerados, existen muchos otros que son de utilidad para cuantificar las variaciones del ADN. Para el lector interesado, recomendamos los trabajos de R. Dickerson y de W. Saenger que se citan en la bibliografía. En condiciones normales de hidratación, la forma predominante de ADN es la B, pero en circunstancias especiales esta forma puede transformarse en A o Z y éstas, a su vez, pueden reconvertirse en B o interconvertirse entre sí. El pasaje de una forma a la otra se hace muy rápidamente, en especial el cambio de B a A, que ocurre en fracciones de segundo. La transformación del ADN B en A se produce en condiciones de moderado descenso de humedad relativa. Por encima del 92% de humedad todo el ADN tiene la forma B, pero tan pronto como la humedad relativa desciende al 80-83 %, comienza la transición en algunas de sus regiones. Finalmente, a un 75% de humedad relativa, la mayor parte de la molécula se encuentra como forma A (sin embargo, aun en estos bajos niveles de humedad aquellas regiones muy ricas en pares de bases AT son capaces de persistir como forma B). Según Dickerson, la caída en los niveles de humedad relativa produciría una deficiencia de moléculas de agua en el surco menor del ADN mientras que persistiría un nivel adecuado de hidratación tanto en el surco mayor como en la cadena de fosfatopentosas. La consecuencia de esta situación sería la pérdida de profundidad del surco menor, el aumento de profundidad del surco mayor y la adopción de la forma A. El pasaje a la forma Z ocurre en presencia de soluciones salinas (por ejemplo de cloruro de sodio o cloruro de magnesio) de alta concentración. Cuando una secuencia repetida de tipo CG (o GC) adopta la forma Z, existe una marcada cercanía entre los fosfatos de las dos cadenas complementarias, y como estos fosfatos poseen cargas negativas tienden a repelerse y producen una resistencia a la formación del ADN Z. Ahora bien, si existen suficientes cationes (cargas positivas) disueltos, éstos se asocian con los fosfatos, neutralizan sus cargas e inducen así la aparición de la forma Z. Es evidente que la célula viva no presenta condiciones de humedad relativa baja o de alta salinidad, por lo tanto, si las formas de ADN A y Z sólo existieran en las circunstancias antes descriptas, tendrían interés biofísico pero carecerían de importancia biológica. Sin embargo, ha sido probado que ambas formas aparecen en la célula viva. Consideremos, por ejemplo, qué es lo que sucede durante el proceso de transcripción de la información genética del ADN al ácido ribonucleico ARN. Tanto el ADN como el ARN reciben su nombre de la pentosa que integra su molécula, la desoxirribosa en el primer caso, la ribosa en el segundo. El carbono en posición 2' de la desoxirribosa se encuentra asociado con dos hidrógenos mientras que en la ribosa está asociado a un hidrógeno y a un oxidrilo, OH. Este grupo OH de la ribosa se encuentra cerca de distintos átomos de los fosfatos y bases de la molécula de ARN produciendo fenómenos de repulsión que obligan al polímero a adoptar la forma A. Durante el proceso de transcripción, una de las dos cadenas complementarias de ADN sirve como molde para la síntesis de una cadena de ARN a la que transfiere su información. En ese momento, la cadena de ADN adquiere la forma A de la cadena de ARN transitoriamente ligada a ella. Es por lo tanto evidente que en toda región cromosómica con actividad transcripcional, el ADN cambiará de B a A, para volver a B una vez que el proceso de transcripción se haya corrido a otra región. Este proceso es catalizado por la enzima ARN-polimerasa, que posee un sitio activo de una longitud equivalente a 40 pares de bases de la molécula de ADN. Así, resulta evidente que la zona donde tiene lugar la transcripción y la consecuente formación de ADN A tendrá una extensión mínima no inferior a 40 pares de bases. Diversos experimentos han demostrado, en los últimos años, la existencia de ADN Z en la célula. La macromolécula de ADN puede adoptar una forma lineal o una forma circular cerrada. Gran parte del ADN de las bacterias y de los virus, el ADN mitocondrial y el de los plásmidos, adoptan formas circulares. Aunque en general se acepta que el ADN nuclear de las células eucariotas (células de los seres superiores con núcleos bien definidos) se halla organizado en largas unidades de cadena abierta o lineal, una importante cantidad de datos experimentales tiende a modificar este concepto. En el núcleo en interfase (período entre dos divisiones celulares) gran parte de la fibrilla de cromatina se halla organizada en forma de múltiples bucles o asas. Los dos extremos de cada una de estas asas se unen a estructuras de la membrana nuclear denominadas complejos de poro nuclear y se comportan, por lo tanto, como una unidad circular cerrada. Dado que cada una de las asas contiene ADN, queda probado que el núcleo de la célula eucariota aloja múltiples unidades de ADN circular. El ADN circular puede encontrarse en forma relajada o en forma superenrollada. En la forma relajada, el círculo se halla desplegado sobre un único plano; en la forma superenrollada el contorno del círculo va girando sobre sí mismo de manera tal que adquiere profundidad (véase la figura 8). Las dos formas de ADN circular pueden visualizarse en el microscopio electrónico como círculos relajados o superenrollados. Además, pueden identificarse por electroforesis o por centrifugación (separación de partículas en suspensión coma ayuda de un campo eléctrico o de un campo gravitacional respectivamente). En estos casos, la estructura compactada del ADN superenrollado aumenta su migración electroforética y su velocidad de sedimentación, lo cual permite diferenciarlo del ADN circular relajado o del ADN lineal. Como casi todas las propiedades físicas, químicas y biológicas del ADN se vinculan al ADN circular y a su estado relajado o superenrollado, la clara definición de estas formas es importante para alcanzar una acabada comprensión de dichas propiedades. La matemática resulta, en estas circunstancias, un aliado indispensable. Dado que el ADN es un polímero con propiedades elásticas,cuando se produce la ruptura de una de las dos cadenas complementarias en una molécula superenrollada, la cadena rota gira sobre la sana hasta que se pierde el superenrollamiento. Por tal motivo, cualquier agente físico (radiaciones ionizantes) o químico (bleomicina, radicales libres, etc.) capaz de romper el ADN tiene la capacidad de relajar la molécula circular. Cuando la ruptura del ADN acontece en la célula viva, ocurre habitualmente un proceso enzimático de reparación que cierra la brecha con rapidez y restituye la integridad del ADN circular. En estas condiciones es factible que se recupere también el superenrollamiento. Las modificaciones del enrollamiento del ADN circular se realizan a través de enzimas denominadas topoisomerasas. La topoisomerasa II tiene la propiedad de transformar un ADN circular relajado en superenrollado. Este proceso implica pasar de una forma sin almacenamiento de energía a una forma con alto contenido energético, y por lo tanto es necesaria la presencia de adenosinatrifosfato (ATP) como donante energético.Por el contrario, la transformación de ADN superenrollado en relajado con subsecuente liberación de energía es catalizada directamente por la topoisomerasa I sin necesidad de ATP. En organismos eucariontes, las topoisomerasas se ubican selectivamente en la membrana nuclear, punto de inserción de las asas de cromatina que forman los dominios circulares de ADN. Esta localización resulta ideal para que la enzima promueva o corrija cambios de superenrollamiento que afectan a toda el asa cromatínica. Como se explica detalladamente en "ADN circular y matemática topológica", una modificación estructural en una región limitada de la molécula circular de ADN modifica la topología alolargo de todo el círculo. Estos cambios arquitectónicos son sumamente importantes porque modifican la relación del ADN con proteínas reguladoras y ejercen, por lo tanto,un profundo efecto en el funcionamiento génico. Ha sido demostrado recientemente que el modelo de Watson y Crick no es el único. En efecto, se han descripto asociaciones dedos, tres y cuatro hebras de ADN que se interconectan mediante uniones no complementarias de guanina. Estas estructuras, denominadas de Hoogsteen en homenaje a quien las describió por primera vez en 1959, se encuentran en los extremos (telómeros) de cada cromosoma eucarionte. Recientemente, nuestro grupo de investigación junto con J. Kere de la Universidad de Helsinki, ha podido demostrar que algunos genes producen múltiples copias de sí mismos que se asocian entre sí y con el ADN de copia única mediante estructuras de Hoogsteen. Lejos ha quedado la época en que se pensaba que una molécula constituida por sólo cuatro nucleótidos diferentes no podía tener la versatilidad y ductilidad necesarias como para almacenar la información genética de un ser vivo. Se pensaba entonces que las proteínas con sus veinte aminoácidos reunían las condiciones adecuadas para actuar como depósitos de genes. Pero hoy sabemos que pese a su limitada variedad química, el ADN puede organizarse y estructurarse en formas muy cambiantes. Esta maravillosa molécula contiene todos los genes de las formas vivas de nuestro planeta, es capaz de autoduplicarse,de transmitir la información genética, de modificar su arquitectura y modificar el funcionamiento génico,de mutar, fijar y también corregirlas mutaciones. Durante los últimos cuarenta años se han develado la estructura química del ADN y el código genético (véase "El lenguaje de la vida". Las próximas décadas permitirán comprender mejor su arquitectura y topología, así como la forma en que microcambios moleculares provocan macrocambios en el funcionamiento génico. LECTURAS SUGERIDAS CRICK,F. H.C.,1976," Liuking Numbers and Nucleosomes", Proceedings of the National Academy of Sciences, U.S.A, 73, págs. 2639-2643. DICKERSON,R.E.et. al.,1982, "The Anatomy of A, B and Z-DNA ", Science 216, págs. 475-485. FREELAND-JUDSON, H.,1980, The Eight Day of Creation, Simon and Schuster, New York. LAFER, E. M. et. al.,1981, "Antibodies to Lefthanded Z-DNA Bind to Interband Regions in Drosophila Politene Chromosomes", Nature 294, págs. 417-422. SAENGER, W., 1984, Principies of Nucleoic Acid Structure, Springer-Verlag, Berlin-Heidelberg. PD: mas imagenes en la Fuente
Hola, mi nombre en Bruno Bianchi, tengo 24 años y soy estudiante de la Licenciatura en Ciencias Biológicas de la Facultad de Ciencias Exactas y Naturales de la Universidad de Buenos Aires (Argentina). Actualmente estoy terminando la carrera y realizando mi tesis de licenciatura. Desde hace unos meses estoy en el Laboratorio de Neurociencia Integrativa trabajando en neurolingüistica. En Febrero de este año lazamos un experimento masivo sobre lectura de cuentos, intentando explicar cómo predecimos las palabras a medida que vamos leyendo un texto. Hoy, después de mucho tiempo, vuelvo a estar comunidad a pedirles que me ayuden completando el experimento. Objetivo del experimento El objetivo del experimento es estimar la predictibilidad de una palabra dado el contexto en el que se encuadra. Para esto vas a tener que completar palabras faltantes en diferentes textos (ver las instrucciones). Siempre se completa con una sola palabra y esa palabra debe ser aquella que te suene más factible según el contexto (poné la primera que se te venga a la cabeza). Hay palabras muy predecibles (muchas personas responden con estas palabras) y palabras muy poco predecibles (las cuales menos del 10% de las personas responde). Eso nos hace diferentes y enriquece al lenguaje, así que no te preocupes si la palabra que completaste no es igual a la original, es parte de la naturaleza que intentamos describir y explicar con este experimento. Instrucciones Durante este experimento deberás leer textos literarios. Cada texto se muestra de a partes, deteniéndose regularmente para que completes con una sola palabra. Luego de completar la palabra deberás apretar ENTER para que el texto continúe hasta la próxima palabra faltante. En cada texto se deben completar alrededor de 100 palabras. Esto no es una competencia con palabras correctas o incorrectas, deberás completar con la palabra que te parezca que va a continuación según el contexto en el que estás leyendo. Tené en cuenta que podés completar con cualquier tipo de palabra, no sólo sustantivos o verbos. Una vez terminado un texto (aproximadamete 20 minutos) podrás cerrar la ventana o seguir con otro texto. En caso de cerrar la ventana podrás volver en otro momento utilizando el mismo mail y seguir desde donde estabas (si cerrás antes de terminar, se pierden los datos, y no podrás completarlo luego). En total son 8 textos, completá todos los que puedas (¡tendrás más chances de ganar el premio! ¡y nosotros más datos!) Premios ¡Cada texto que completás suma 100 puntos para el sorteo (Si no completás el texto hasta el final, los puntos no se suman)! Podés completar hasta 8 textos. Y si completás todos, ¡te damos 500 puntos extra! Mientras más puntos tengas, más chances de ganar alguna de las órdenes de compra* ( Click aquí para ver los ganadores hasta el momento). Recordá que cada texto se debe completar en una única sesión. Podés cerrar la sesión, volver a entrar con el mismo mail cuando quieras y seguir completando otros textos, pero si dejaste un texto incompleto no podrás hacerlo luego.
Médicos fumadores: Haz lo que yo digo... En Argentina los médicos fuman sólo un poco menos que el resto de nosotros. En países desarrollados los médicos dan el ejemplo y fuman mucho menos. En nuestro país mueren aproximadamente 45000 personas cada año por enfermedades relacionadas con el consumo de tabaco en todas sus formas. El trabajo de investigadores argentinos muestra que la capacitación de los médicos es clave para mejorar su posición respecto al tabaquismo y los consejos que brindan a sus pacientes. Horacio M. Zylbersztejn, Alberto Cardone, Nora Vainstein, Andrés Mulassi, José G. Calderón, Patricia Blanco, Enrique Pautasso, Aníbal Picarel, Román Cragnolino, Susana Fernández, Adriana Andina, Sebastián Saravia Toledo, Italo Torchio y César A. Belziti, junto a un importante número de colaboradores, realizaron una encuesta sobre 6497 médicos de todo el país. El estudio busca información sobre el hábito de consumo de tabaco entre los médicos y el impacto sobre sus pacientes. Tabaquismo: Haz lo que yo digo...Tabaquismo: Haz lo que yo digo... El trabajo de Zylbersztejn y colaboradores muestra que alrededor del 30% de los médicos fuma. En la población general este porcentaje es algo mayor (entre el 33 y el 39%). Sin embargo en países desarrollados los médicos fuman mucho menos que la población general. En Holanda, por ejemplo, el 40% de la población fuma, pero sólo el 6% de los médicos consume tabaco. El estudio hecho en Argentina destaca que los médicos que recibieron entrenamiento sobre el tratamiento del tabaquismo tienden a fumar menos ellos mismos y a dar consejos más a menudo a sus pacientes fumadores para ayudarlos a dejar el hábito. Algunos datos interesantes extraídos de la encuesta se refieren a la gran variación del porcentaje de fumadores por especialidad. Los médicos que trabajan en emergencia son mucho más propensos a fumar (21% más). Esto puede deberse al estrés asociado a este tipo de trabajo. Por otro lado, los cardiólogos y neumonólogos fuman proporcionalmente menos que los médicos del resto de las especialidades. Se especula que esto se debe a que los especialistas de corazón y pulmones son los que comprenden más claramente el efecto del humo del cigarrillo en el organismo. Curiosamente, los médicos que ejercen especialidades quirúrgicas fuman más que los médicos de otras especialidades, aún cuando algunos de estos pueden ver directamente los efectos devastadores del tabaco al operar a sus pacientes. Uno de los resultados más significativos del análisis, que confirma estudios hechos en la población general, es que los hijos de padres fumadores o exfumadores tienen hasta cuatro veces más posibilidades de volverse fumadores. Esto muestra cuan importante es el ejemplo de los padres hacia los hijos. Los investigadores insisten en que formar a los médicos argentinos en el área de tabaquismo es fundamental. Sólo un tercio de los médicos encuestados había tomado algún curso o asistido a charlas informales sobre tabaquismo. Los que así lo habían hecho recomendaban más frecuentemente a sus pacientes tratamientos para dejar de fumar. Con una inversión mínima en capacitación podrían obtenerse importantes mejoras en la salud pública. Trabajo original: Tabaquismo en médicos de la República Argentina. Estudio TAMARA, Revista Argentina de Cardiología , vol 75, pp 109-116 (2007) Instituciones: Consejo de Epidemiología y Prevención Cardiovascular y por el Área de Investigación de la Sociedad Argentina de Cardiología .

Registrate y eliminá la publicidad! Bueno, leyendo un poco el post sobre el canon, se me ocurrio hacer una busqueda sobre campañas anti-canon para hacer una pequeña campaña anti canon en esta comunidad... bueno, para empezar me gusto este video español que explica un poco el dilema que hay con el canon...o sea..si pago los derechos de autor tengo derecho a usar los objetos que compre para llenarlos de cosas registradas link: http://www.videos-star.com/watch.php?video=mkCR8T0ujn8 la misma asociacion que hizo el video de arriva nos deja esta parodia a coca-cola link: http://www.videos-star.com/watch.php?video=M5ywlwyVmNY&eurl esto es un video de CQC españa en contra de SGAE (la asociacion que se encarga de cobrar el canon en españa) link: http://www.videos-star.com/watch.php?video=9BHc-bcBZXs&rel=1 y aca les dejo este pequeño debate español entre un abogado que se opone al canon y miembros de la SGAE link: http://www.videos-star.com/watch.php?video=QsBw3n803v4&eurl siguiendo, aca encontre un ejemplo de lo que pasa con una familia española normal que tiene una computadora...cuanto gasta al año en canon digital...despues explica un poco lo que gana SGAE con la venta de computadoras al año Cita :Una familia normal en España con: - 1 ordenador con 160gb de disco duro (22E de canon) - 1 regrabadora de dvd’s de ordenador (16,67E de canon) - 1 impresora multifunción (10E de canon) - 1 cámara de fotos con memoria para 200 fotos (9E canon) - 1 reproductor de dvd de salón (6,61 E de canon) - 1 Equipo de música de salón (0,60 e canon) - 1 reproductor de mp3 (14e canon) - 1 línea adsl 1 MB (35e canon “anuales”) - 200 cd’s vírgenes para grabar diversos datos (50e de canon) - 100 dvd’s vírgenes para grabar diversos datos (140e de canon) En definitiva, que cualquier familia española con un ordenador paga unos *303e de canon al año*. Antes no se sabia que era lo del canon, pero ahora nos vamos a enterar de sobra, por lo que veo. Seguimos con los datos: 234,5 millones de ordenadores que se venderán en 2006 (previsión de vunet),lo que hace un total de: *710,5 billones de euros de canon* (casi nada). ¿Y esto para los cantantes? Y esto nada mas que de ordenadores domésticos. Ni que pensar de los ordenadores portátiles y ordenadores que se venderán a empresas que también tendrán estos cánones y su uso no será para grabar música. fuente:http://www.intruso.info/2007/03/27/campana-anti-canon-sgae/ Estos son los afiches españoles contra el canon terminando por ahora voy a dejar post sobre el canon que han aparecido en T! para empezar, el post que lo explica todo (por pichicha) http://taringa.net/posts/info/1046045/%C2%BFQue-es-el-canon-y-porque-deber%C3%ADa-importarte-Muy-claro.html luego pichicha nos deja un listado de musicos a favor http://taringa.net/posts/info/1046263/Quienes-se-adhieren-a-la-famosa-_Equot;Ley-Nacional-de-la-Mu.html Cabezon19 da 5 razones para no aceptar el canon http://www.taringa.net/posts/offtopic/1049599/Cinco-argumentos-para-decirle-NO-al-canon-digital.html Pabloneuquen nos explica un poco algunas formas de lograr que no acepten la ley http://www.taringa.net/posts/noticias/1057061/C%C3%B3mo-oponernos-a-la-Ley-del-Canon.html aca les dejo la pagina de la asociacion "no al canon" www.noalcanon.org Algunas direcciones para hacer saber tu opinión: info@musicosconvocados.com legales@musicosconvocados.com gremiales@musicosconvocados.com federal@musicosconvocados.com culturales@musicosconvocados.com prensaydifusion@musicosconvocados.com eventos@musicosconvocados.com cmorgado@diputados.gov.ar Acá algunos banners que se realizaron: YAPA: buano para finalizar les dejo un problemita que tuvo la pagina de la SGAE por si no se entiende bien paso a explicar...el logo de la campaña española contra el canon aparecio en la pagina de SGAE, esto fue resultado de un bug del sitio fuente: http://n0xtrum.blogspot.com/2007/04/campaa-anticanon-de-la-sgae.html Bueno por ahora esto es todo...por mas info les deje los post de otros usuarios, si encuentro mas info o uds me la pasas la agregare...esto me costro trabajo asi que tratemos de no bardear mucho..su comentario constructivo se agradece <a href='http://b.t.net.ar/www/delivery/ck.php?n=a2afc290&cb=INSERT_RANDOM_NUMBER_HERE' target='_blank'><img src='http://b.t.net.ar/www/delivery/avw.php?zoneid=58&cb=INSERT_RANDOM_NUMBER_HERE&n=a2afc290' border='0' alt='' /></a>